東京大学教養学部 駒場総合科目D
森林生態圏の保全と管理 第4回講義 生物社会としての森林(動物) 石田 健 1999.11.02
前書き
「森林生態圏」という用語は、講師=石田にとっては天から降って湧いたようなもので、造語した人物から説明を受けたこともなく「なんのこっちゃと」いう気持ちである・・・だから、学生諸君と同じである。ここはひとまず前向きに取り組み、あえてこの用語の意味を見いだそうとすれば、生物間相互作用と物質循環の(無機環境を含めた一定の=ある仮定のもとに、閉じた)系を想定した「生態系」にその外部である人間の社会(活動)との「物質および情報の出し入れ」を加えた仮想空間(時間を含めた4時限と考える)を、生態圏とよぶ・・・ことにする。その保全というのは、少し古い森林学用語を使えば、「持続的森林経営」のことであり、それと森林生態系に焦点をあてた管理、がこのオムニバス形式の科目の共通??テーマ(縦糸)でしょう・・・たぶん。
私の担当は、動物。森林に存在する物質としての生命体においては、樹木が圧倒的な量を占めており動物などは微々たる量でしかない。たとえば温帯林で、1ヘクタールあたり樹木の300トンに対して、大小全部合わせた動物は数キロというところで、10の5乗の差がある。木材の大部分は死んだ組織だが、生きた部分だけでも10の3乗ていどの差はゆうにある。
ひるがえって、現在記載されている150万種余りの地球生物の内、100万種余りが甲虫を中心とした昆虫・節足動物が占めていて、植物は30万種足らずである。地球生命は未知のものを含めると数千万種から1億種くらいいると推定されているが、その大半もやはり昆虫なのである。昆虫は、1個体は小さいものの個体数も種数も大きいので、重さで評価するとたとえば熱帯のアリだけで大型のヒョウ一頭分にもあたるそうだ。森林におけるアリなども、いろいろな話があって面白い。
私は鳥や獣の生態を軸に、森林を研究している。森林における量においても種数においても優占しない生物の話が、単なる嗜好を超えて森林生態圏の保全と管理にとってどう重要なのか、講義の終わりまでに多少とも納得してもらえたら、私の目的は果たされる。もともと興味のある学生には、気軽に楽しんで聴いてもらえればよい。
キーワード:森林、masting=種子の非定常的生産、野生鳥獣、生物間(動植物の)相互作用、進化論、適応
本論の序文
近年では、一部の新聞でディアウォーズ(シカ戦争)などという言葉が出てきたり、尾瀬でハイカーがツキノワグマに襲われたというような話しがけっこうよく報道される。人間は、森林を自分の都合によいように使おうとするが、自然相手ではそうは問屋がおろさない。森林には、野生鳥獣が生息しており、それらも森林を自分に都合のよいように利用しようとする。そこで、人間との利害関係が生じる。そして人間にもさまざまな生き方がある。近年、森林資源が減少し、一方で人口が急増しつい先日60億人を超えたそうだ。地球上での人間と野生動物、人間どうしの利害関係の対立がますます顕在化している。
シカ(ニホンジカ、エゾシカ)やカモシカは、1950年後半からの一時期には天然林の伐採によって絶滅を危惧する人もいたほど減少していたが、造林地=彼らにとっての餌場の増加が主因になって各地で個体数が増加した。カモシカはなわばりを持つので密度増加が先にとまったものの、シカは非常に高密度になり、人工林のみならず自然植生の破壊者としても懸念されている。ニホンザルは、主に林縁部の農作物への加害、ツキノワグマやヒグマは農業被害の他に、クマハギという植林木への加害やときには人身傷害事件なども発生する。これらの問題は、端的に言えば、野生動物の存在を軽視してきたツケが出てきているのであるが、森林生態圏の管理上は社会的な利益とコスト(経費・労力)のつりあいを見定める必要もある。
これらの、人間対野生動物、人間対人間の2つの局面において利害の調整をどうするか。調整の方途を照らすために、事実を明確にする必要がある。例えば、各地域での個体数やその動向、個体数の変動を左右する要因を明らかにし、また動物の生態・行動の特質をよく理解した上で、実効のある対策を検討することが肝心だろう。本講義では、ブナ科樹木のマスティングという現象に焦点をあて、それが野生動物の生態とどう関わっているかを示す。実例として、このような性質をもった樹木種子を主要食物とするツキノワグマの、着床遅延や非常に小さい仔の出産、冬ごもり、植物を主食とする食性などの生態的特徴と森林動態との関係について簡単に紹介する。
表1.秩父山地における堅果の豊凶記録
-----------------------------------------─
年 ブナ イヌブナ ミズナラ* クリ**
-----------------------------------------─
1984 豊作? 凶作
1985 凶作 凶作
1986 豊作? 凶作
1987 凶作 凶作
1988 凶作 豊作
1989 豊作? 凶作
1990 並作? 凶作
1991 凶作 凶作
1992 凶作 凶作 凶作 凶作
1993 並作 豊作 豊作 並作
1994 凶作 凶作 凶作 豊作
1995 豊作 凶作 並作 並作
1996 凶作 並作 凶作 並作
1997 凶作 凶作 並作 並作
1998 凶作 凶作 豊作 並作
1999 凶作 凶作 並作? 豊作
-----------------------------------------─
大久保ら(1989)、梶ら(1992)、梶ら(未発表)、石田・
橋本(未発表)の資料による。
*ミズナラは個体差が大きく、並作は結実量の異な
る木が混在した年。
**クリと1999年は、目視観察による質的評価。
マスティング
日本の冷温帯(落葉広葉樹)林の代表樹種であるブナは、一定しない数年から10年近い間隔をあけて堅果を大量に成らす(表1)。太平洋側山地帯に生育するイヌブナもほぼ同様であり、ミズナラなどいわゆるドングリを成らすナラ・カシ類もブナ属ほど顕著ではないが、非定常的な種子生産をする。その他の樹種も、毎年一定の結実をみるものはむしろ少ない。これらの種子は栄養価が高く厳しい冬を前にした秋に大量に得られることから、多くの動物が好んで食べる。例えば、秩父山地での調査で、1988年にイヌブナの樹下で1平方メートルあたり約950個、1993年にブナの樹下で同じく約990個の種が落ちてきた。ブナやイヌブナの種(殻を除いた中味だけ)は約0.3gあり1000個は0.3kgである。つまり、1平方メートルあたり300g、1ヘクタールあたりだと種だけで約3トンの種が落ちたことになる。樹上で鳥や小型ほ乳類などの食べられてしまった種もあるので生産量はそれ以上。ブナやイヌブナが森林にまんべんなく密生しているわけではないが、膨大な量であることは理解される。凶作年には、まったく結実しなかったり、1998年にはブナが樹上ではかなり多く結実したが、地上に落ちるまでにほとんどすべてが昆虫に食べられて中味がなかったという場合もあった。
マスティングは、亜寒帯の針葉樹から熱帯の常緑樹まで多くの樹種で知られている。例えば、1996年に日本の研究者が多く参加している熱帯林研究プロジェクトの調査地、マレー半島のランビルで樹木の一斉開花が起こった。熱帯では、昆虫によって花粉が媒介される樹木が多いが、それでも、樹種が多く多様な熱帯林では開花が同調することは重要な意味を持つ。また、花粉を媒介する昆虫をめぐる競争もあるだろう。
マスティングが進化した原因
マスティング現象は、その樹木にとって適応的で、自然淘汰によって進化してきた考えられる。進化の原因として、種子捕食者飽和(回避)説、風媒説、環境予測説、資源マッチング説、動物媒促進説、動物散布促進説などがあげられている。
マスティングがみられる樹種の多くは、大きな栄養価のある種子をつける。大きな種子は、発芽時に成長のためのエネルギーを多く持つことが自身にとっても有利であると同時に、重いこと自体は種子散布に不利である。このような種子は多くの動物を引きつけ動物によって母樹から離れた場所へと散布される。
結実量に大きな変動があり、しかも変動周期が一定しないと凶作のときには種子食者を飢えさせ個体数を抑制することができ、凶作に続いて豊作があると、その急激な量の変化に種子食者の数的増加は追いつかず多くの種子や実生が残存できる。この仮説は、理屈としてはよくできているし、動植物の種間関係についてより一般的な原則を議論することのできる魅力的な仮説である。
多くの樹種は、風媒花で、多くの樹種は自家受粉では種ができなかったり発芽率が悪い自家不和合性を持ち、開花・結実は地域全体で同調させる。そのような場合には、特に熱帯林のように同一樹種が近隣に極めて少ない状況では、多くの花を集中して開花させることが、受粉を確実にする。
大きな種子を多量につけることは母樹にとってかなりの負担を伴うと推定され、毎年行うことは難しいかもしれない。条件がよい時に行うことは有利であり、また豊作の後には必然的に凶作が続くかもしれない。ただし、大きな種子を大量に生産することは、母樹にとってはかなりのコストかもしれないし、実はたいしたコストではないのかもしれない。ブナのように寿命が300年ていどもある木にとっては、数年から10年ていどの豊凶の周期も、一生のごく短いできごとかもしれない。
マスティングが進化した原因を説明しようとする仮説は、互いに排他的な関係ではないし、いくつかの原因が、それぞれの地域の環境やそれぞれの樹種の特性および進化の歴史に応じて関わっているのが事実かもしれないし、進化の過程で重要だった要因が今も同様に重要だったとは限らない。
ツキノワグマの生態
ツキノワグマの食性
ツキノワグマは春先には高茎草本を、その後には草木の葉を、夏には葉に加えて漿果や無脊椎動物を、そして秋には堅果を中心に食べる。その間に交尾、妊娠、冬ごもりという3つの大きなできごとがある(図1)。
ツキノワグマの食物となる広葉樹の葉や果実は、質と量のいずれにおいても季節的な変化が大きい。量的に集中して出現する食物は、春には新葉であり秋には堅果である。春の新葉は、ツキノワグマの生息地である落葉広葉樹林においては毎年一定して大量に生じ、ツキノワグマにとっては年変動はないも同然である。一方、秋の堅果は、種ごとにみるならば非常に大きな年変動があることが知られている(表1)。
橋本(1995)が糞分析した結果、ブナとミズナラ(ドングリ)が同時に豊作だった1993年秋には、主にミズナラを食べており、ブナを食べる必要がなかったようである。一方、ミズナラとブナとイヌブナが不作だった1994年の秋には、クリが豊作となり、食物はほとんどがクリであった。このように、堅果を成らす樹種が多種共存している森林では、全部の堅果が凶作となるいわばツキノワグマにとって飢饉の年は、むしろ少ないのかもしれない。
ツキノワグマはたいへん広食性で、あるものはほとんどなんでも食べているように思われる。食肉目の消化器官をそのままひきつぎながら、植物質にかたよった食性をもっているので、脂質や蛋白質を比較的多く含む堅果類の消化率は高いとしても、多くの食物の消化効率は悪いと推定される。ツキノワグマは、大きな体を維持するためにはかなり大量に食べる必要があるはずだ。
そのために、堅果類(と蜂蜜?)は特別としても、ツキノワグマの食べ物の構成は、主に摂食効率によって決まっているのではないかとも予想される。ツキノワグマの生態をより深く理解する上で、利用可能な食物資源の空間的時間的分布と、食物資源にたいするツキノワグマの採食行動の対応を詳しく調査することが、今後の重要な研究課題の1つだと言える。
ツキノワグマの採食行動の特徴として、大きな体に似合わず木登りが上手で、樹上の資源を有効に利用できる点も重要であろう。それによって、同じ食物資源でも樹上と地上の両方で採食できるために、利用可能な期間が伸び、利用できる食物の種類が増える。森林という高さのある空間を立体的に利用することによって、大型獣としてはきわめてコンパクトな環境利用様式をとれることは、ツキノワグマの強みであろう。
ツキノワグマの環境選択
ツキノワグマは、おおざっぱにいえば食物のあるところにいる。大きなスケールでは、年間を通して食物が確保でき、少なくとも近隣で、たまには、秋の堅果が十分に得られる植生域に生息(定着)している。ツキノワグマにとって価値の高い、柔らかい葉や果実を多く産するのは広葉樹林であり、なかでも落葉広葉樹林が主な生息環境である。
ツキノワグマは森林性ではあるが、その中での植生選択の幅は広く、現在の分布域をもっとも強く規定しているのは、人間の活動だと言える。分布域が人間活動に強く規定されていることは、逆に、ツキノワグマと人間の環境選択の類似性と過去における競争関係を類推させる。行動圏を調べると、ツキノワグマは人間の生活圏にも居るごく身近な動物であることがわかる。
採食空間としての環境選択および人間活動の影響は、比較的定着性の高い雌と大きな遊動域を持つ雄では、かなり違いがあるはずだ。
雌の生活史特性
研究上の評価基準としては、ある個体が一生の間にどれだけ多くの子供を残し、そのうちのどれだけ多くの子供が繁殖できるようにまでなるか(生涯適応度)、といったことが一般的には問われる。ツキノワグマの成獣にとっては、いかに多くの食物を得られるか、それも秋に堅果を十分食べられるかが生涯適応度を左右していると推測される。
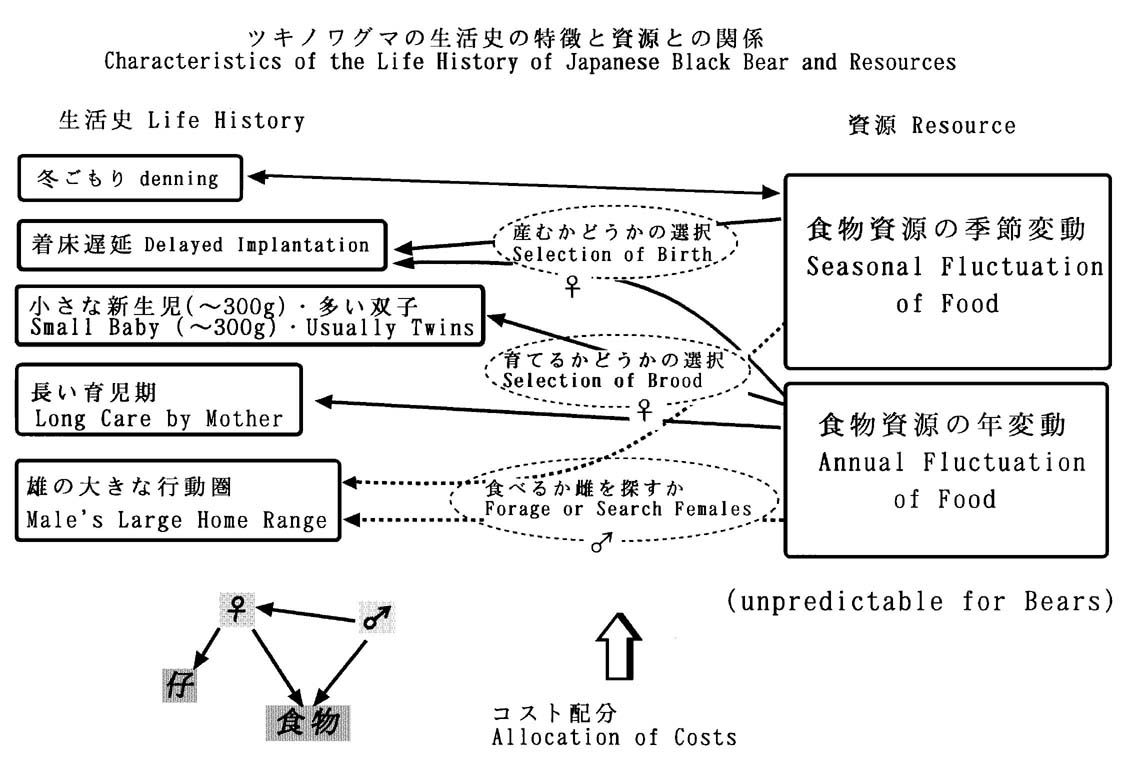
大人の雌グマは、初夏に交尾をしたあと、受精卵を子宮に着床させずに保持し、秋の終わりに妊娠する。野生のツキノワグマにおいて直接実証されているわけではないが、間接的な資料から、クマ類に有名なこの着床遅延の生理特性を、本種も持っていることは間違いない。受精卵を持っている雌は、秋にせっせと採食し、十分な栄養が得られれば妊娠し冬ごもりの間にきわめて小さい仔を産む(図2)。産仔数は2頭が多いらしい。
妊娠後、どこまで仔の世話をするか、あるいはできるかはそのときどきの食物条件の結果としての母グマと仔グマの生理状態によって決まると仮定できる。母グマの立場からは仔の面倒をより多く見て仔の生存率を高めるべきか、自分自身の摂食を増やして次の繁殖に備え、より長く生きるべきか、損得勘定しているとも比喩できる。仔の成長がよければ早く手放しても仔の生存率は高くなるという要素や、双子で1頭だけ育つか2頭育つかといった微調整もしているという要素も考えうる。
雌は、交尾期も採食に専念し、夏の間も徐々に体重を増しているらしい。この時期には、仔をもっている雌が仔の保護(育児)を続けて仔に投資するか、仔別れして雄と交尾するかといった、選択肢が考えられる。
クマの仔が親の体重比できわめて小さいこと、クマの重要な捕食者が(人間をのぞいて)ほとんどいないことから、1回ごとの妊娠や仔の世話の負担や繁殖失敗による損失は、雌グマにとっては比較的軽くてすむと言える。一方、消化率の低い、変動する食物で大きな体を維持しなければならないわけで、自分自身の将来と仔への投資の損得勘定においては、前者により重きがおかれていると推定される。
したがって、ツキノワグマの雌の生活史特性を要約すると、常に自身の摂食を中心に生きており、初夏に交尾・受精をしておいた上で、妊娠するか、仔の面倒を見続けるか、そのときどきの餌条件によって、柔軟に対応していると言える。重要な食物である堅果の量は大きく年変動するが、数年待てば必ず豊作の年がある。繁殖負担が軽く食物条件に柔軟に対応できる性質は、理にかなったものに思える。
また、秩父においても、雌は、比較的一定し、雄に比べて小さい行動圏(平均的に7~8平方キロ)を持つ。先に、堅果樹種が複数共存する森林では、堅果結実量の年変動が少ないようだと述べたが、定着性の強い雌においては、区域内での樹種構成の相違によって個体間での生涯適応度に差が生じるであろう。雌の行動圏のありかたも、食物資源の分布や変動に左右されている可能性が高いが、雌の定着過程や個体の入れ替わり、間接的な個体間競争(雌雄でも)、生涯における行動圏の変化などについては未解明である。
雄の生活史特性
雄は、大きな行動圏を持つ。秩父では、最外郭法で評価して最大60平方キロの範囲を1年間に動いた個体と、食物を求めて直線距離で約9km移動した個体が確認された。雄は、初夏には雌グマを、春と秋には食物を中心に活動している。秩父で、再捕獲された同じ個体の体重変化を検討すると、直接の記録は1個体についてだけだが、雄の成獣は夏の間に大きく体重を減らしている。この期間は、採食活動を切り詰めて雌を追い求めていると言える。
雌が、食物と仔との3角関係で生きているのに対して、雄は食物と雌との3角関係で生きている。食物がより豊富な春と秋には採食に重きがおかれ、そのときに蓄えた脂肪を糧にして夏には雌を追って、活発に移動しているらしい。雄は、夏の間だけ、個体維持のために採食したり余り動き回らずにエネルギーを節約するか、交尾するために動きまわるかの損得勘定をする。
雄の体重の変化は大きく、20~30%の増減が記録された。雌も、雄ほどではないが10~20%の変化があった。冬ごもり直前の体重も含めれば、雌雄とも40%前後も体重が増減していると推定される。ツキノワグマは移動することと耐えることの2つの手段によって、季節と年ごとに変動する食物資源に対処している。移動量は雄が雌よりも大きく、雌においては、着床遅延後の妊娠の有無がそれまでに体に蓄積できた栄養量によって決められていると考えられ、個体維持という点では第3の手段を持っているとも言える。雌雄の生活史戦略の差異は、雄が動くことに偏った戦略を持っているのに対して、雌は耐えることに偏った戦略をとっているとも言えよう。
マスティングとツキノワグマと人間
マスティングを説明する説の1つとして、動物による種子補食活動があげられ、すべてではないにしても有力な仮説として研究や議論が行われている。
ツキノグマは、ブナやミズナラにとってはおそらくそれほど重要な種子補食者ではない。別の動物が規定した森林の性質を受けて、その生態が独特に進化している。クマにとっても、20年近い一生の間に確実に仔を残せるならば、数年以上の不確定な間隔での結実もそれほど難しい問題ではない。
キノワグマによる農林業被害、ときには人身生涯事件などに対してどのような対処が必要だろうか。
考慮すべき重要な点は、(1)森林にクマは「いるもの」であるという認識を持つこと、(2)クマのような動物には個体差(個性と言っても良い)が大きい、(3)動物は、基本的に個体が遺伝子を残すことに有利な行動をとる、という3点を理解することだろう。現在の社会的コンセンサスとしてクマの地域個体群を絶滅させることなくかつ農林・人身被害を防止するには、地域ごとのクマの動態・行動を把握し、一定の緩衝地帯を設けて、クマを人間の活動から遠ざける対策が必要だろう。
石田のホームページ:研究の一部を紹介してあります。
http://forester.uf.a.u-tokyo.ac.jp/~ishiken/ken.html
または
(http://www.uf.a.u-tokyo.ac.jp/Chichibu/person/ishida/japanese/ken.html)
図1.ツキノワグマの食性と年周活動
http://forester.uf.a.u-tokyo.ac.jp/~ishiken/graphics/kuma/lifecycle.jpg
に掲示
図2.ツキノワグマの生活史特性
http://forester.uf.a.u-tokyo.ac.jp/~ishiken/graphics/kuma/lifstyle.schema.jpg
に掲示
オマケの図 ブナとミズナラの種子トラップと実
http://forester.uf.a.u-tokyo.ac.jp/~ishiken/graphics/masting/seedtrap.nut.acorn.jpg
に掲示
参考資料(授業のときは省略)
Dave Kelly. 1994. The evolutionary ecology of mast seeding. Trends in Ecology and Evolution. Vol.9 No.12 [102]:465-470.
predator satiation, wind pollnation, environmenal prediction, resource matching, animal pollination, animal dispersal accessory costs, large seed size ---hypotheses for masting
湯本貴和.
1999. 熱帯雨林. 岩波新書 岩波書店,
東京
本体 740円 1999年7月19日発行
{kind=link}
{kind=link}
{kind=link}