BIRDER 1999(9) 1999.7.14
飛ばない翼の進化
─ 飛ぶべきか飛ばざるべきかの”自然選択”
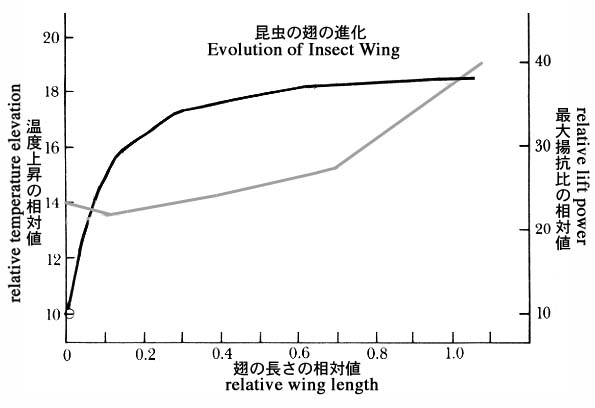
『鳥のように空を飛んでいきたい』と人は言う.飛ぶことは鳥のいちばんの特徴だし,飛ぶための翼や羽根こそ鳥に独特のものだと誰しもまず思いつくだろう.飛ぶための翼は,生物の進化を考えるときに,目や脳などとならんでよくひきあいにだされる器官である.進化のしくみや化石の証拠にもとづくならば,完璧に飛べるような翼があるとき突然出現したのではないと,明らかに判断される.それでは,初めの内は飛ぶことに役にたたなかったような小さな翼から,飛ぶための翼のような構造が自然選択されて徐々に進化してきたと考えてもよいのだろうかという,疑問が指摘されダーウィン以来多くの進化学者は答えに窮してきた.この疑問に対する現代の標準的な解答は,翼は最初は飛ぶことに適応して進化したのではなく,体温調節など別の機能が自然選択されて翼が大きくなるように進化し,あるときから飛ぶことに転用されたのだというものだ.翼が小さい内は翼の面積の増加にともなって熱交換機能が速やかに増加し,翼が一定の面積を超えると熱交換機能はほとんど増えなくなり,代わりに空中で揚力を生む機能が増え出すということが,昆虫サイズ・昆虫型の翅では実験的に証明されている(図1).
鳥は昆虫よりもかなり重く,鳥の翼は昆虫のものより大きく,羽根で形成されているので性質もかなり異なり,この図をもって鳥の翼の進化にそのままあてはめることはできない.しかし,鳥の翼の進化でも,同じように,機能の転換が起こったと推測する参考になる.翼が小さい内は熱交換や保温,少し大きくなってからは虫取り網代わりや走るときにバランスをとったり滑空したりする道具として役立ち,自然選択され大きくなったのだろうという説がある.現生の鳥も,つがい形成の儀式行動,卵やひなの日陰や傘,影をつくって魚をおびき寄せるなど,飛ぶ以外の多くの用途に翼を利用している.
かくして鳥は空を飛ぶ能力を獲得した.しかし,翼や飛ぶための筋肉の成長,あるいはそれらを使って飛ぶために,鳥はたいへん大きなエネルギーを使っている.つまり,鳥が飛ぶということは,飛ぶ必然性がそうとうに高い,あるいは飛ぶ能力が強い自然選択圧を受け続けていると考えられる.したがって節約できるものなら,その分のエネルギーを産卵数を増やしたりヒナの世話をすることにまわして多くの子孫を残した個体の方が有利なはずである.環境しだいで,飛べない翼が自然選択されるということだってあってしかるべきなのだ.実際に,ダチョウのように陸地を走りまわったり,ペンギンのように水中を泳ぐことのほうが得意な飛ばない鳥もまた多い.「鳥類の起源と進化」という本の中で,A・フェドゥシャ(1996)は,本文のほぼ6分の1を飛ばない鳥の章にさいている.現在生きている飛ばない鳥たちは,すべて,飛んでいた祖先の鳥から進化した鳥たちだという.飛ばない鳥や飛ぶのが不得意な鳥がどのように進化し適応しているかを知ることも,鳥やその翼のことをよく理解する上で大切なのである.
飛ばない鳥の多くは,大型である.翼の面積は長さの二乗の関係で増減し,筋肉や骨の体積に比例する体重やエネルギー消費量は基本的に長さの三乗の関係で増減するので,大きな鳥は相対的に飛ぶことの負担が大きい.一方,大きな鳥ほど,自分より大きな捕食者が少ないので飛べないことの不利が小さかったり,主に長さの二乗の関数である体表面積に影響される摩擦抵抗と長さの三乗の関数である体重に影響される重力の関係で,水中を泳ぐのに有利だといったこともその原因である.
アフリカ大陸のダチョウ,南アメリカ大陸のレア,オーストラリアのエミューなどが代表例で,ニュージーランドにはモアという巨鳥がごく最近まで生存していたし,今も体の割に巨大な卵を産む数種のキーウィがいる.飛ばないで泳ぐことを得意とする南半球のペンギン類は,ヒレのように進化した翼で水中を飛ぶように移動する.北半球には,最近までペンギンに似た別系統のオオウミガラスがいた.一方で,泳ぐときには翼をしっかりたたんで脚ヒレで泳ぐのがアビのなかまであり,その一部の鳥は飛ばない.
そして,飛ばない鳥は,島に多い.島に飛べない鳥がいることには2つの意味がある.1つは,島が飛べない鳥にとって最近まで安全地帯だったこと.大昔には大陸や海にもいた飛ばない鳥の多くが,競争相手や捕食者のほ乳類に駆逐されてしまい一部しか残っていないという歴史的意味である.ヨーロッパや北アメリカ大陸の暁(ぎょう)新世と始新世の化石で知られる史上最大の鳥ダイアトリマや,南アメリカ大陸のより最近の漸新世から鮮新世にかけて生息していた
phorusrhacid (Phorusrhacos, Brontornis, Andalgaloris・・・)が知られている絶滅鳥である.南半球の巨大ペンギンや,北半球の日本からも化石が見つかっているペンギンに形の似たペリカン目に近縁の体長が2mもある巨大海鳥(Plotopteridae)がいて,どちらも中新世のころに絶滅している.
もう1つは,島では飛ぶことへの投資を節約することが,有利になるという自然選択の意味である.島には,渡って来られなかったり狭くて住み着けなかったりして,飛べないほ乳類の競争相手や捕食者がほとんどいなかったから,飛べないことが淘汰されず,飛ばずに効率よく繁殖する性質が選択されてきたのである.
ガラパゴス諸島のガラパゴスコバネウ,ニュージーランドの小島に生残するオウムの一種であるカカポ,ニューカレドニアにいる独特のカグーなどが代表例である.そしてグループとして例数の多いのが,ヤンバルクイナを始めとするクイナ科の中型の鳥たちである.島嶼性のクイナの約4分の1が飛ばないとい言う.
一方,スズメ目を初めとした小型の鳥では,知られている飛ばない種の例数はわずかで,「灯台守の飼っていた一匹の猫に滅ぼされた」という逸話(ほんとうはそう単純ではなさそう)で有名なニュージーランドのスティーブンイワサザイもその一例である.また,ほとんど飛べなかったホオジロ属の「絶滅した新種」の遺骨が,カナリア諸島の洞窟から見つかったと,アメリカ合衆国の鳥学雑誌に最近報告された(図2).
飛ばない鳥は,島では最近までもっと多かったと想像される.近世になってから記録にあるスズメ目の3種が絶滅種として記録されており,モーリシャス島のドードーやニュージーランドのモア,ハワイ諸島のハワイガンなどの大型種も加えると,島で絶滅した飛ばない鳥の数はかなりの数にのぼる.とりもなおさず人類が,島を飛べない鳥の安全地帯でなくしてしまったのである.
島の鳥には,自然選択の上でもう一つ考慮すべき事情があると,私は考えている.島が大陸やとなりの島から遠い場合には,その島へやってくるのに強い飛翔力が必要だろう.住み着いた最初の個体が強い飛翔力を持っていれば,その形質はあるていどその子孫にも遺伝すると推定される.しかし,島に住み着いた後には,飛べなくならないまでも大陸にいたときほど強い飛翔力を必要とせず,飛ぶ形質への選択圧は弱いと言える.多くの種の若鳥は巣立った場所から分散するので,風が強い島では,むしろ,飛翔力が強く遠くへ飛ぼうとする個体は海の藻屑と消えてしまう危険性が高く,淘汰されやすいという場合もあると考えらている.前に,大きな負担のある飛ぶという性質は,強い選択圧があって初めて維持されるだろうという趣旨のことを述べた.その強い選択圧が弱まれば,いずれは飛ばない方向への進化が起こると推定される.
すると,島にたどりつくには飛翔力が必要で,島にいついてからは飛翔力がないほうが有利になる.この矛盾した傾向が,実際に島に生息している多くの鳥ではどうなっているのだろうか.進化は,人生の長さにくらべるとはるかに長い時間をかけてゆっくりと起こる.飛ぶ能力を維持する選択圧が減って,飛べない翼が進化していくにしても,それにはそれなりの時間が必要である.しかし,こうした進化の事実を観察するの好都合な鳥をみつけて,詳細に観ていくと,ダーウィンフィンチのくちばしのように,進化の断片らしきものが観察できるということが最近になって例示されつつある(ワイナー
199 ).島の鳥の進化を観るためには,あるいていどの数の島に分布してきたくらいのほどほどの飛翔力や分散する性質を持っていて,かつ,自然選択の方向が変わったときにそれに応じて変化が観られるていどの飛翔力の遺伝的変異を持っていて,かつ,島への分布が起こってから識別できるていどの進化の起こった時間の経過している種,が最適だろう.
コゲラは,日本列島周辺に広く分布し多くの小さな島々にも生息している.少し意外なことに,佐渡島や伊豆大島などの比較的「本土」に近くあるていどの面積もあり森林もあるいくつかの島に生息していない.とても遠くて小さい小笠原諸島や南西諸島の先島列島などにいないのは納得できるとしても,近くの島にいないことは少し謎めいていないだろうか.私は,このことと,コゲラが1980年代になってから突然のように東京などの都市緑地で繁殖し始めたこととは,コゲラの性質と関連した似た現象なのではないかという仮説をたてた.都市緑地は,小さな緑の島に例えることのできる特質を備えている.1980年代は,その緑の島がそれ以前よりも緑の本土である山地帯から遠くなった時代でもあったと言えるだろう.このようなコゲラを研究することは,島の鳥の翼の進化のことを知るよい道しるべになるのではないかと期待できる.
図3は,コゲラの右の翼の初列風切羽の長さを,外から2番目から始めて内側に向かって1枚ずつ7枚測定した値を,本州で捕獲した個体と奄美大島で捕獲した個体とで比較した結果である.奄美大島のコゲラの翼はやや短く,先端が丸い傾向がある.同じ体重の鳥であれば,翼が小さいほど飛ぶのに不利であり,近縁種どうしや種内では長距離の渡りをする集団の方が,短距離しか季節移動をしない集団の鳥よりも翼が尖っている傾向があることが報告されている(Monkkonen
1995).沖縄島のコゲラの翼はもうわずかに本州のコゲラの翼に近いものの短く,三宅島のコゲラは本州のものにもっと近いという結果になっている.残念ながら,まだまだずっと多くの計測値を得て比較しなければダーウィンフィンチにおいて観察されたような進化の様子をかいま見ることはできないし,私の仮説の可否についてはっきりとした判断を下すことはできない.
参考文献:Feduccia(1996)The Origin and Evolution of Birds, 420pp.;Rando&Lopez(1999)Condor 101, 1-; Monkkonen,M.(1995)Evol. Ecol.9,520-, ワイナー『フィンチのくちばし』(19 )早川書房,
コゲラの死体を拾得された方は,研究室(〒113-8657 東京大学・院・農生命・森圏管理学研)までお送りいただければ幸いです.(送料は,石田が負担いたします)
http://forester.uf.a.u-tokyo.ac.jp/~ishiken/ e-mail:ishiken@uf.a.u-tokyo.ac.jp
図説明文
図1.昆虫サイズの昆虫型の翅をアルミ箔などで模したモデルによって,翼の面積と熱交換機能および飛翔力(揚力)の関係をしめした実験結果を要約した図.翼が小さいうちには,その面積の増加にともなって熱交換力がどんどんますが,その機能はやがて頭打ちになる.熱交換機能が頭打ちになったあたりの翼面積からこんどは揚力を生む機能が急激に大きくなって飛ぶための翼としての価値が生まれている(グールド
19 の図を改変).鳥類の翼は羽毛によって形成されており,もっとずっと大きいので両者の関係はこの結果と同じではないが,同様の機能変換が進化の途上で起こったと推定する参考になる.
図2.アシナガホオジロの想像図(Rando&Lopez 1999を参考に描き直す)
図3.コゲラの初列風切の1枚ずつの長さの変化を,本州のコゲラと奄美大島のコゲラで比較した図.繁殖前期(営巣前の時期)につがいを形成していた野生の繁殖個体の成鳥のみの測定値を比較した.まだ資料が不十分だが,奄美大島のコゲラの翼のほうが丸い傾向がみられる. 参考図,本州と沖縄島の場合の比較
図4.コゲラの翼先端部と初列風切長の測定部位の例.いちばん外側の初列風切は,極端に短く,相対的な長さも一定しない.外側から7枚目以降で急速に短くなる.
島にすむ動物は中型化するというのが,いちばん一般的な傾向である.島というのは小さい陸地なので,そこにすめる生き物の個体数にも種数にも多様性にも限界がある.大陸でなら,似たような生態を持っていても大きな種と小さい種で資源をわけることができるが,島では1種でぜんぶ使わないと生きていけないというような事情のせいだと言われている.小さい動物は大型化する.私が体重測定をしたコゲラの中でいちばん重かったのは,奄美大島にいたメスであった.伊豆諸島のメジロはくちばしが長い,奄美大島のオーストンオオアカゲラは南方にもかかわらず他の亜種より大きいなどの例がある.
{kind=link}
.jpg){kind=link}
-2.jpg){kind=link}